花了不止 15 分钟的时间,整理了一下分子生物学中名字里有“子”的概念。(本来只是想看一下 Khan Academy 生物 AP 里 U6L5 的内容,结果又加上了《Essential Cell Biology》、Nick Lane 的《The Vital Question》和 Phillip Ball 的《How Life Works》的一些内容)
先讲两个内容相互独立的小段子,请勿恶意联想。
北大学霸来到山河四省的一所高中介绍学习经验。
学霸在台上问大家:“我能考上北大,最大的秘诀就是勤奋!你们凌晨六点都在干什么?”
下面答:“刚上完一节早自习,咋了?”
学霸扶了一下眼镜:“所以你们的问题就在于不懂得劳逸结合~”
理查德·费曼 (Richard Feynman) 当时想了解一点生物学,于是去旁听一门生物课。老师让学生轮流做报告。
费曼对生物学几乎是外行,轮到他做报告时,他觉得必须从基础讲起,于是:
- 在黑板上画了一只猫的轮廓
- 开始标出各个肌肉的名字
结果还没讲多久,下面的生物学生就打断他说:
“We know all that!”(这些我们早就知道了!)
费曼就回了一句非常经典的话:“那难怪我这么容易就追上你们了——你们学了四年生物。”
然后他补了一句更毒的评价:他们把时间都浪费在记这些东西上,而这些 15 分钟查书就能找到。
以上段子和本文内容无关——
粒子物理中,在各种探测器中可以留下轨迹的对象往往被命名为“XX子”,英文名的词尾都是 -on,比如 photon, electron, lepton, proton, neutron, hadron, baryon, meson…
分子生物学中,基因表达过程众多参与者的汉语名也往往是“XX子”,但是——
- 相应英语的词尾有的是 -on,有的是 -er/-or;
- 前面的词根有各种顾名难以思义的近义词,比如“启动/增强/激活/引导”、“沉默/抑制/阻遏”;
- 之所以要用这么多近义词,主要是为了区分同一个过程中,参与者的不同化学本质:DNA 区域、蛋白质、小分子……
——合理怀疑是东施效颦效到一半绷不住了。
花了不止 15 分钟的时间,整理了一下分子生物学中名字里有“子”的概念。(本来只是想看一下 Khan Academy 生物 AP 里 U6L5 的内容,结果又加上了《Essential Cell Biology》、Nick Lane 的《The Vital Question》和 Phillip Ball 的《How Life Works》的一些内容)
演员表:
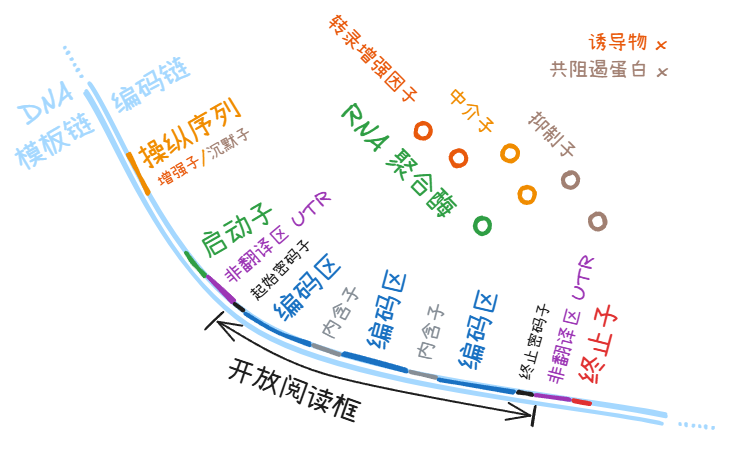
剧目表:
DNA 区域
转座子 (transposon)
转座子也被称为“跳跃基因”。
有些 DNA 分子上的区域,可以在生物的生长过程中把自己移动到 DNA 的其他位置,甚至是其他的 DNA 上。也就是说,基因在染色体上的位置不是固定的。
这些可移动的 DNA 区域就被称为转座子 (transposon)。
操纵子 (operon)
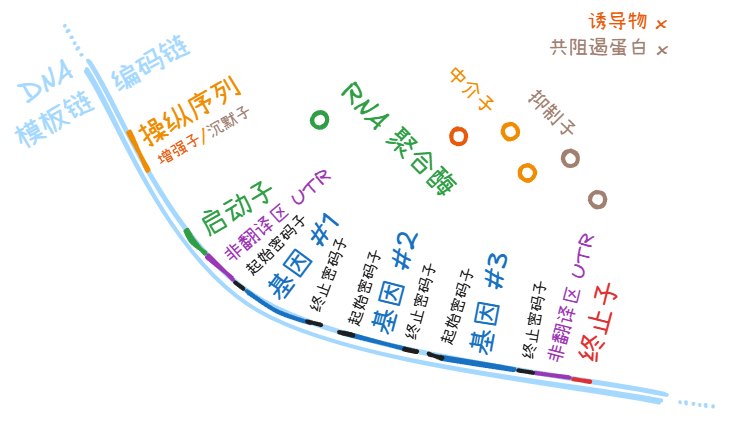
操纵子 (operon) 指的是原核生物的一个启动子 (promoter)、 该启动子相关的操纵序列 (operator)、控制的下游的多个基因组成的一大段序列。此概念的关键是多个基因转录在一条 mRNA 上。
操纵子 (operon) 是一个主要存在于原核细胞的概念,在真核细胞里极少(不是没有)。
但是那些自带一套启动子、操纵序列的基因,看成只有一个基因的操纵子也……
注意把操纵子 (operon) 和下面的操纵序列 (operator) 区分开, 操纵序列 (operator) 是操纵子 (operon) 的组成部分。
调控序列 (regulatory sequence)
操纵序列 (operator)
是操纵子 (operon) 的一部分,和转录因子 (transcription factor) 结合,可以改变启动子 (promoter) 和 RNA 聚合酶结合的效率。
刚刚发现操纵子 (operon) 的时候,理论认为操纵序列 (operator) 位于启动子和基因之间,与抑制子 (repressor) 结合,阻断 RNA 聚合酶向 DNA 下游滑动。
后来才发现有些操纵序列 (operator) 和转录因子 (TF) 提高转录水平,有些降低;所处的位置也不固定;调控的机制也是多种多样……细节越来越丰富,全景越来越模糊。
真核生物既然很少谈 operon,也就往往不谈 operator,而是说“增强子 (enhancer)/沉默子 (silencer)”。
转录因子结合位点 (TF binding site)
单个结合位点一般比较小,长度只有十几到几十个碱基,但是结合位点附近的 DNA 区域对于转录都有调控作用,下文里的增强子 (enhancer)、沉默子 (silencer) 等等往往一个区域内包括好几个转录因子结合位点**。**
有些调控区域距离启动子 (promoter) 在基因序列上的间距非常大,甚至中间相隔若干其他基因。因为 DNA 分子在细胞中盘曲折叠,DNA 链上相隔很多碱基的两段序列,在物理空间中反倒可能因为折叠而紧邻对方。
增强子 (enhancer)
= DNA 分子上转录激活因子 (activator) 的结合位点附近的区域。(见“转录因子”一节)
沉默子 (silencer)
= DNA 分子上抑制子 (repressor) 的结合位点附近的区域。(见“转录因子”一节)
绝缘子 (insulator)
略。(怎么越整理越多啊)
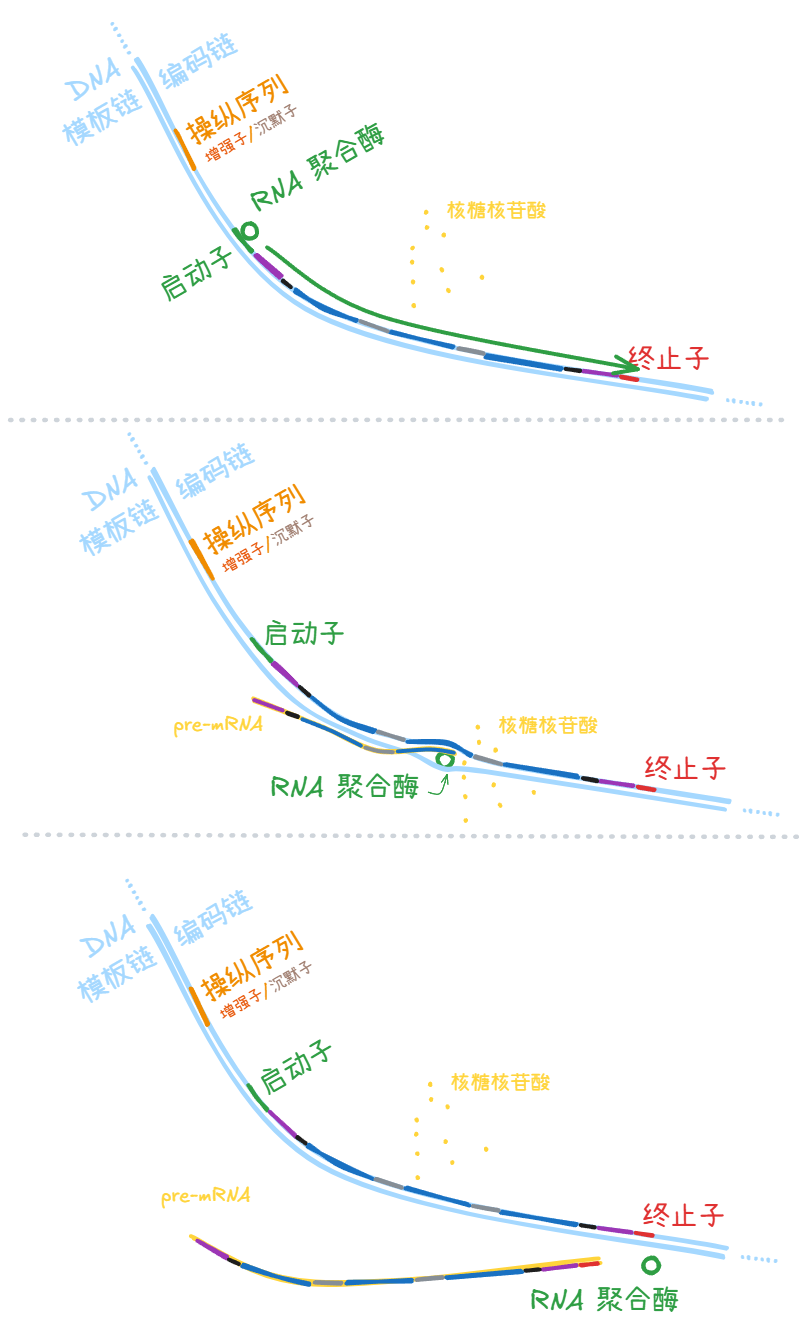
启动子 (promoter)
= RNA 聚合酶在 DNA 分子上的结合区域。
所谓的“启动”特指转录的启动。
细胞中游离的 RNA 聚合酶先结合到启动子 (promoter) 上,然后从模板链的 3’ 向 5’ 方向滑动,滑动过程中 RNA 聚合酶附近的 DNA 双链解旋,游离的核糖核苷酸与模板链的碱基互补配对,聚合成一条和编码链信息相同的单链 mRNA 分子。
中文没有问题,问题在于 promote 这个单词一般语境下不会翻译成“启动”。
终止子 (terminator)
= RNA 聚合酶在 DNA 分子上的脱钩区域
启动子 (promoter) 和终止子 (terminator) 的一个区别是:启动子 (promoter) 一般不会被转录,而 RNA 聚合酶是在终止子 (terminator) 被转录完了才脱离 DNA 分子,所以终止子 (terminator) 的序列信息会出现在转录成果,也就是 pre-mRNA 中。
DNA/RNA 区域
本节内容的概念,其序列信息既可以出现在 DNA 分子上,也可以出现在 RNA 分子上。(前面的终止子 (terminator) 也是,但是为了和启动子 (promoter) 对偶就没放在这里。)
就像是同样一篇文章,既可以用毛笔写在绢帛上,也可以用铅笔写在白纸上,但是绢、纸、水墨和石墨中的原子和分子们本身并不是文章。
信息的载体和信息本身,并不是一回事。
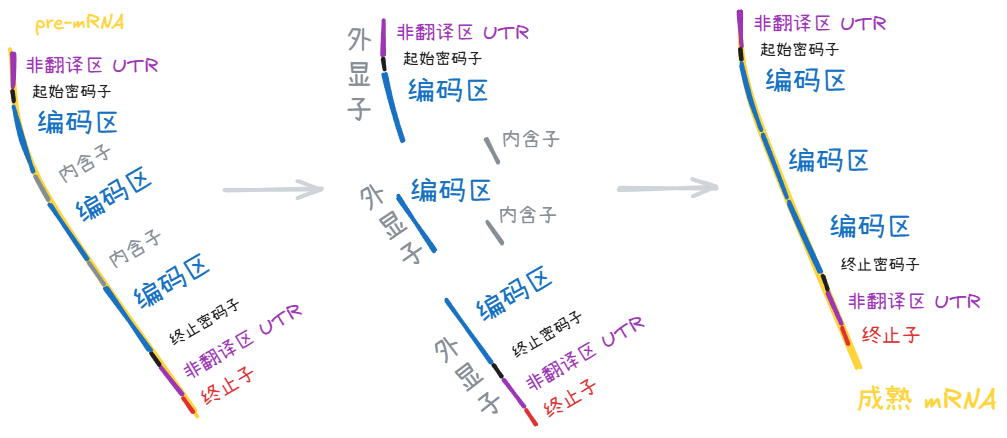
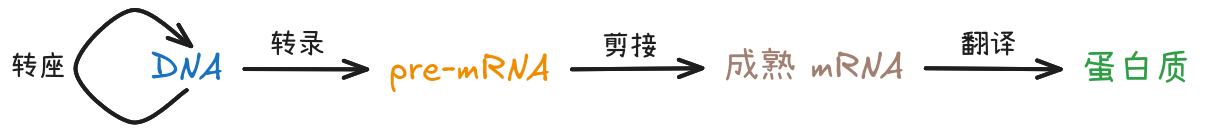
本节的各位演员们主要靠两个过程定义自己的身份:转录后修饰、翻译。
真核生物转录 (transcription) 生成的 pre-mRNA 链,其中的内含子 (intron) 被切除,剩下的区域连接在一起——这一过程叫 RNA 剪切 (splicing)。剪切前后 mRNA 分子的头尾再做些其他修饰,变成成熟的 mRNA 分子。
成熟的 mRNA 和核糖体结合后,核糖体向 mRNA 分子下游滑动的过程中,游离的氨基酸被链接成多肽链——这一过程叫翻译 (translation)。
非翻译区 (UTR)
转录是从启动子开始,到终止子为止;
然后 mRNA 接受转录后修饰;
翻译从起始密码子开始,到终止密码子为止。
mRNA 上 5’ 端起始密码子之前,3’ 端终止密码子之后,各有一部分并没有被修饰掉,但其中的信息也不会进入下游的蛋白质,称作非翻译区 (UTR)。
开放阅读框 (open reading frame, ORF)
DNA 或 RNA 上,从起始密码子 (start codon) 开始,到终止密码子 (stop codon) 为止的连续核酸序列。
密码子 (codon)
每 1 个氨基酸通过 3 个连续核苷酸编码,这 3 个核苷酸的碱基种类序列称作 1 个密码子。
起始密码子 (start codon)
核糖体和 mRNA 分子结合后,翻译开始的位点,其本身也编码一个氨基酸。
终止密码子 (stop codon)
翻译结束的位点,本身不编码氨基酸,而是让核糖体和多肽链脱离 mRNA 分子。
编码区 (coding sequence, CDS) 和非编码区
所谓的“编码”特指编码蛋白质。也就是最终决定蛋白质氨基酸序列的那部分 DNA/RNA 序列。
非编码区作为编码区的补集,范围可就大了,内含子、非翻译区、调控序列,甚至基因之间的不明意义的区域都是。
内含子 (intron)
mRNA 剪切过程中被去除的部分。
外显子 (exon)
RNA 剪切过程留下的部分。
由于内含子这个概念更本位,外显子是作为内含子在 mRNA 序列上的补集而存在的,所以会出现一些很拧巴的命题:
非翻译区属于外显子。
外显子中的序列信息并不全都“显示”在蛋白质中。
……
蛋白质
以转录 (transcription) 调控为主线,蛋白质为主角,各种“子”之间的结合关系可以列成下表:
| DNA 区域 | 蛋白质 | 小分子 |
|---|---|---|
| 启动子 (promoter) | RNA 聚合酶 (polymerase) | |
| 操纵序列 (operator) | 转录因子 (transcription factor, TF) | |
| 增强子 (enhancer) | 转录激活因子 (activator) | 诱导物 (inducer) |
| 沉默子 (silencer) | 抑制子 (repressor) | 共阻遏物 (corepressor) |
| 中介体 (mediator) |
RNA 聚合酶 (polymerase)
催化转录过程,和启动子 (promoter) 结合后,沿 DNA 分子的模板链滑动,方向是从模板链的 3’ 到 5’ 端,所以转录出来的 mRNA 和 DNA 编码链 5’ 到 3’ 端的信息相同。
转录因子 (transcription factor, TF)
与操纵序列 (operator) 结合以后,可以改变转录效率。早期发现的转录因子主要通过影响 启动子 (promoter) 和 RNA 聚合酶 的结合效率来发挥作用;后来发现的工作机制越来越多,比如chromatin remodeling, mediator recruitment, enhancer-promoter looping 等等……
一般用 Hill 方程来建模,受调控基因的表达速率 v
- [S] 是游离的转录因子的浓度,
- 是最大表达速率,
- 参数等于达到最大速率一半时的转录因子浓度,
- n 是 Hill 系数,表示转录因子的调控方向和结合位点的合作程度。
![Hill 系数分别为 +1(橙色)、+5(红色)、-5(蓝色)时的 v([S]) 函数图像,其余参数都取 1。](/photos/2026-06-14-hill-equation.png)
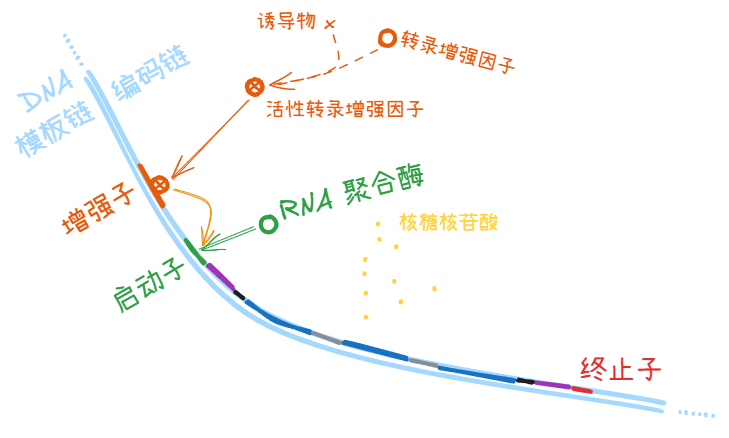
转录激活因子 (activator)
和 DNA 分子上的增强子 (enhancer) 结合,提高转录效率。
此时的 Hill 系数 n > 0 (图中红线)
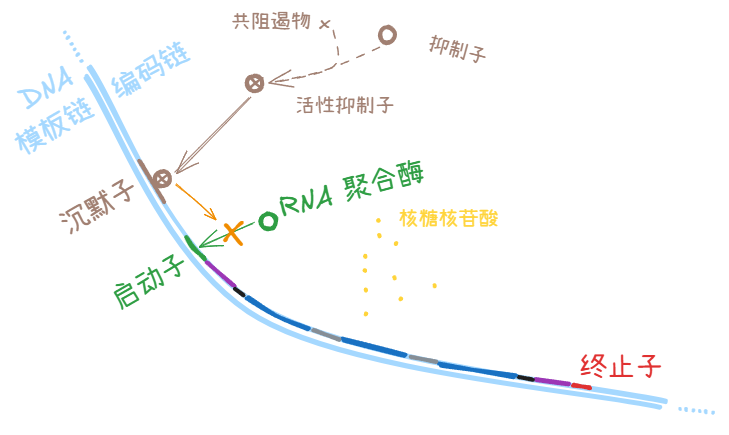
抑制子 (repressor)
和 DNA 分子上的沉默子 (silencer) 结合,降低转录效率。
此时的 Hill 系数 n < 0 (图中蓝线)
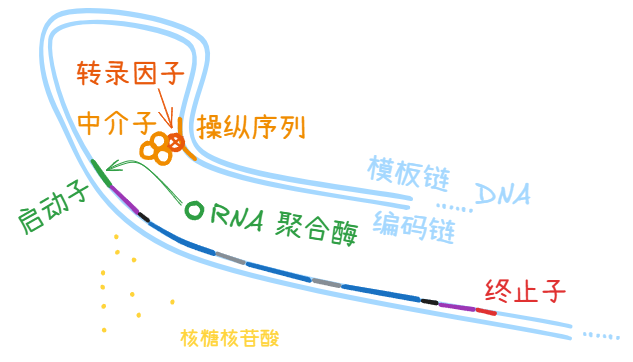
中介体 (mediator)
怎么不叫中介子了?
前面说过,有些转录因子 (transcription factor) 距离启动子 (promoter) 在基因序列上的间距非常大,靠着 DNA 分子在细胞中盘曲折叠,才在物理空间中靠近。
有时这个距离对于扒在增强子 (enhancer) 上的转录激活因子 (activator) 来说依然不够近,因此需要和一定数量的中介体 (mediator) 形成复合体,才能够到启动子 (promoter),吸引 RNA 聚合酶前来结合。
小分子
所谓“小”是相对于蛋白质和 DNA 分子的原子量,这些分子不需要对细胞特殊处理,就可以从细胞外环境进入细胞内部。
这些小分子可以和转录因子 (TF) 结合,改变转录因子 (TF) 的结构,进而影响转录因子 (TF) 和操纵序列 (operator) 的结合效率,让人可以在细胞外主动改变细胞内某些基因的表达程度。
因为转录因子有促进和抑制两种调控转录的方向,这些小分子有些促进又有些抑制转录因子的活性,所以一共有 4 种情况,但是往往只按最终效果将小分子分为 2 类:
诱导物 (inducer)
向细胞培养液里添加一些 诱导物 (inducer),就可以让细胞在一定时间内表达出更多下游被对应 转录激活因子 (activator) 调控的基因产物。
有些诱导物 (inducer) 和转录激活因子 (activator) 结合,使后者转入活跃状态,以便和 DNA 上的增强子 (enhancer) 结合;
另外一些诱导物 (inducer) 和抑制子 (repressor) 结合,使后者转入不活跃状态,难以和 DNA 上的沉默子 (silencer) 结合。
共阻遏物 (corepressor)
向细胞培养液里添加一些共阻遏物 (corepressor),就可以让细胞在一定时间内暂停/更少地表达下游被对应抑制子 (repressor) 调控的基因产物。
有些共阻遏物 (corepressor) 和抑制子 (repressor) 结合,使后者转入活跃状态,以便和 DNA 上的沉默子 (silencer) 结合。
另外一些共阻遏物 (corepressor) 和转录激活因子 (activator) 结合,使后者转入不活跃状态,难以和 DNA 上的增强子 (enhancer) 结合。
写得不好,概念太乱了,不如随便看一本分子生物学课本,从原核细胞干到真核细胞,从 DNA 复制一路看到翻译。
世界是一个巨大的草台班子,我是人证,这篇文章是物证。
本文收录于以下合集: